秦岭典型松栎混交林群落种间关系研究
摘 要:研究了秦岭地区群落物种的生态位宽度,生态位重叠指数和种间联结关系,该地区的优势种为油松(Pinus tabuliformis Carr.)、锐齿栎(Quercus aliena Bl. var. acuteserrata.)、白檀(Symplocos paniculate.)和忍冬(Lonicera japonica Thunb.)等,其中油松和锐齿栎在群落中表显出显著的优势度,有较大的生态位宽度,在检验群落中的种间联结性时这些植物总能和其他植物形成较强的正联结关系。群落植物之间存在正联结关系的同时也存在负联结关系,在检验木本植物的种间联结性时,负联结关系多于正联结,说明植物之间存在一定程度的竞争,该群落并未发展到稳定的顶级群落。值得关注的是群落中检测出较大生态位重叠且有较显著联结性的9对种间关系几乎全都发生在重要值较高的物种之间,这说明该地区的优势物种有较强的共存性,研究它们的共存性对该地区群落的恢复和重建有十分重要的指导意义。
关键词:植物群落;种间关系;种间联结性;生态位宽度;生态位重叠
Study on the interspecific relationship of typical mixed forest of pine and oak in Qinling
Abstract: The niche breadth, niche overlap index and interspecies association of plant community in the Qinling mountains were studied. The dominant species in this area are Pinus tabuliformis Carr., Quercus aliena Bl. var. acuteserrata., Symplocos paniculate., Lonicera japonica Thunb., ect. Among them, Pinus tabulaeformis and Quercus acudentatus showed significant dominance in the community with a large niche breadth. When it tests the interspecific association in the community, these plants can always be tested to form a strong positive association with other plants. There are both positive and negative association between community plants, when examining the interspecific association of woody plants, Negative association is more than positive association, it shows that there is a certain degree of competition between plants and the community did not develop to a stable top one. It is worth noting that almost all of the 9 pairs with large niche overlap and significant association are among the species with high importance, it shows that the dominant species in this area have strong coexistence. It is very important to study their coexistence for community restoration and reconstruction in this area.
Key words:Plant community; interspecific relationship; interspecific association; niche breadth; niche overlap
目 录
1 材料与方法………………………………………………………………………5
1.1 研究区域情况………………………………………………………………5
1.2 调查与研究方法…………………………………………………………5
2 结果与分析……………………………………………………………………6
2.1物种重要值和生态位宽度 ……………………………………………………6
2.2 种间联结性…………………………………………………………8
3 讨论……………………………………………………………………………13
参考文献……………………………………………………………………………15
致 谢………………………………………………………………………………17
0 前言
自然界中不同种类的植物生活在一起时往往会与周围的其他植物产生联系[1],比如在资源匮乏时的竞争,一种物种依附于另一物种的寄生关系(比如桑寄生植物的寄生现象),以及有些植物特有的他感作用等,各种各样的联系使处于同一空间的不同物种之间紧密相连,这些关系协同环境的作用共同决定着群落中物种应占据的位置,促使群落朝着特定的方向发展[2],植物之间这种或有益,或有害,或中性的相互作用称为植物的种间关系。目前为止国内外已有很多学者对种群的种间关系进行了多样的研究,其中,植物之间的联结性是学者们十分关注的群落指标。植物的种间联结关系(interspecific association)是指在一个空间中异种植物之间在布局上的相互关联性,这种关联性可能是为正的也可能是为负的[3],正联结通常意味着关联的物种可能共生或存在依附关系,或者对生存环境的要求十分接近;而出现负联结的两个物种则很可能是存在强烈的竞争而相互排斥或者对环境因子的需求大相径庭[4]。植物种间联结关系体现着植物群落的数量特征和结构特点,同时也蕴藏着群落演替过程中种群更替的动态变化[2],结合群落中物种生态位的分析和生态位重叠度的分析,研究植物的种间联结关系有利于我们去理解群落演替过程中物种的形成,探讨群落中物种的以何种方式共存,并且可以在植物群落的修复和重建等过程中提供有效的理论基础。
目前已存在多种对物种种间联结性的分析方法,对群落发展到顶级阶段物种间的联结关系到底该如何发展,不同的学者也有不同的看法,最为普遍的说法有两种:一是认为随着群落越来越稳定物种间的联结性会逐渐趋向无联结的方向发展,二是认为群落越稳定,物种间的正联结性会越来越稳定。不同的说法各有佐证,目前为止哪一种说法更为权威暂无定论[5]。
秦岭是我国中部东西走向的巨大山系,东西跨度达1600千米左右,主峰太白山的海拔高达3771.2米,山势绵延不绝,起伏不定,地形险峻多变。作为我国长江流域和黄河流域的分界线,正处于亚热带与暖温带的交界线上,由于其多变的地形、独特的地理位置和特有的气候条件,秦岭孕育着丰富的物种资源[6],是生态学的天然研究所。秦岭地区的辽东栎是秦岭的地域性顶级群落物种,在一定条件下能形成纯林,人们为了使森林的生产力得到有效发挥和提高森林郁闭度,人为地向辽东栎林中飞播油松,使油松在辽东栎林中自然生长,因此当地存在大部分辽东栎和油松的混交林。本文中的研究地点选在秦岭陕南地区的旬阳坝和火地塘。本论文将通过计算物种的种间联结性,结合物种的生态位和物种之间生态位重叠程度来讨论该地区的群落种间关系。
1 材料与方法
1.1 研究区域情况
本次研究对象为秦岭中段的松栎林,火地塘和旬阳坝都位于陕宁县内,旬阳坝处于都地势自南向北由高降低,平均海拔高约1300米左右,属于北亚热带气候,夏季降水充足,冬季严寒。境内植被覆盖度较高,达到97%,森林资源丰富,物种繁多,松、杉等高大乔木为该地区的主要物种,此外该地区还有多种国家一级珍贵保护树种;该地区也是我国中药的重要产地,盛产天麻(Gastrodia elata)、猪苓(Polyporus umbellatus)、党参Codonopsis pilosula)、五味子(Schisandra chinensis)等中药材[5]。我们在旬阳坝地区不同海拔随机选取九个样点设置一个20m×20m的正方形样地进行群落中乔木的物种信息采集,并在每个20m×20m的大样方随机设置两个2m×2m的小样方进行群落中灌木的物种调查和两个1m×1m的草本样方进行草本植物的调查。调查时记录调查地点的编号、地理坐标、海拔、坡向和坡度,样方中植物的种类、数量、树木胸径、树高、冠幅、盖度等基本信息。
1.2 调查与研究方法
1.2.1重要物种的选择
物种的重要值反映了物种在群落里的相对优势度,因此,想了解一个物种在群落种的地位,就要计算该物种的重要值,同时,群落里出现频率不高的物种其存在有很大可能的偶然性,因此首先要将所有物种的重要值(IV)计算出来,以每个物种的重要值作为该物种在群落中优势度体现的指标,选取重要值较大的种物种进行讨论分析。重要值的计算公式如下[7]:
1.2.2物种生态位和生态位重叠
生态位通常指一个物种在多维空间上占据的位置,体现着物种对资源的利用情况,这里采用Levins于1968年提出,后又经人加权修正的公式计算物种的生态位宽度[8]:
这个公式以物种的重要值作为参照指标,其中,,为物种i在某一资源位j上的重要值,则表示该物种在所有资源位上的重要值之和。假设一个物种的生存范围很广,在所有的资源位中都能被调查出,则这个公式的分母就会变小,得出的生态位宽度也就越大。
计算生态位重叠情况用的是Pianka生态位重叠指数[9]:
和分别表示两个不同的物种i和k在同一样方j的重要值。得出的数值越大,两个物种之间的生态位重叠程度就越大。
1.2.3群落联结性的计算
运用方差比率法[10]预估群落整体的总联结性,这个公式先假设所有选中的物种中不存在关联,根据公式计算检验统计量,公式如下:
其中N为调查的样方总数,ni为调查到物种i的样方数,Tj表示样方j中调查到的物种数,t表示每个样方中调查的物种数量的均值。
当独立性零假设的条件下(即物种之间不存在显著关联时),VR=1。若计算得到的结果VR>1,则说明物种间整体的联结性为正,若VR<1,则群落中物种整体表现为负联结。
种对间的联结性通过计算Pearson相关系数来检验。Pearson相关系数是一种既能反映是否存在联结性又能反应联结性大小的检验成对物种种间联结性的方法。先将需要检验联结性的物种数据按照2×2连列表的方式排列成一个矩阵,再将这些数据代入公式中计算,就能得出相关系数[11]。公式如下:
式中的N为所调查的样方数量,和分别表示一个物种i和另一个物种k在同一个样方j中的密度(多度值),和则表示物种k和物种i在所调查的全部样方中的平均密度。的取值区间为[-1,1],如果取得的数值落在[-1,0),则表示物种i和物种k之间的为负联结,反之则为正联结。
2 结果与分析
2.1 物种重要值和生态位宽度
通过计算物种的重要值,选出乔木中重要值大于1.5的10种乔木,灌木中重要值大于2的11个物种,草本中重要值大于1.5的7种,一共28种植物进行分析,得到的各个物种在对应层次的重要值和生态位宽度的结果见表1到表3所示。
乔木层物种中重要值最大的是锐齿栎和油松,它们的生态位宽度也是最宽的。其余物种重要值和生态位宽度大体上也一致,但在灌木层和草本层的情况就比较复杂,重要值高的物种未必生态位宽度就更广,这说明两者之间并不是绝对对等的,必须将结合起来看,才能得出更客观的答案。
从生态位重叠指数的结果来看,乔灌草三个层次的植物都存在较大的生态位重叠,其中乔木层物种间生态位重叠指数大于0.5的有22对,达到所有种对的48.9%,物种间生态位重叠程度最高,灌木层有14对,占总体的25.5%,草本只有4对,占总体的19.0%。乔木中的油松、锐齿栎,灌木中的悬钩子、桦叶荚蒾、白檀,卫矛,草本植物中的唐松草、苔草、禾本科植物等重要值和生态位宽度较大的一些物种与其他的物种之间的生态位重叠指数较高等,分析这些种对的种间联结关系,发现种间联结性较大的物种生态位重叠也普遍较明显。
表1 乔木层主要物种的重要值和生态位宽度 | |||||
Table 1 Important values and niche width of main species in tree layer | |||||
物种名 | 相对多度 | 相对盖度 | 相对频度 | 重要值 | 生态位宽度 |
Species name | Relative abundance | Relative coverage | Relative frequency | Important value | Niche breadth |
鹅耳栎 | 0.0112 | 0.0027 | 0.0353 | 1.6402 | 2.9941 |
华北落叶松 | 0.0179 | 0.0242 | 0.0039 | 1.5367 | 1.0001 |
华山松 | 0.02915 | 0.0493 | 0.0353 | 3.798 | 2.0650 |
苦木 | 0.0538 | 0.0530 | 0.0353 | 4.7366 | 2.9666 |
梾木 | 0.0493 | 0.0051 | 0.0980 | 5.0816 | 4.4023 |
麻栎 | 0.03812 | 0.0203 | 0.0353 | 3.1228 | 2.3690 |
漆树 | 0.0135 | 0.0122 | 0.0980 | 4.1239 | 4.9095 |
锐齿栎 | 0.4036 | 0.3335 | 0.2510 | 32.9365 | 7.7719 |
油松 | 0.2937 | 0.4519 | 0.2519 | 33.2193 | 7.9454 |
灯台树 | 0.0112 | 0.0032 | 0.0353 | 1.6558 | 2.9994 |
表2 灌木层主要树种重要值和生态位宽度 | ||||||||||
Table 2 Important values and niche width of main species in shrub layer | ||||||||||
物种名 | 相对多度 | 相对显著度 | 相对频度 | 重要值 | 生态位宽度 | |||||
Species name | Relative abundance | Relative coverage | Relative frequency | Important value | Niche breadth | |||||
菝葜 | 0.0972 | 0.0156 | 0.14286 | 8.5229 | 4.5505 | |||||
白檀 | 0.0648 | 0.3024 | 0.0914 | 15.2886 | 3.3236 | |||||
勾儿茶 | 0.0093 | 0.0693 | 0.0057 | 2.8103 | 1 | |||||
胡颓子 | 0.0046 | 0.0670 | 0.0057 | 2.5767 | 1 | |||||
胡枝子 | 0.0417 | 0.0112 | 0.0514 | 3.4755 | 2.5824 | |||||
桦叶荚蒾 | 0.0602 | 0.0220 | 0.0229 | 3.5002 | 1.9147 | |||||
忍冬 | 0.1111 | 0.1458 | 0.1429 | 13.3241 | 3.8499 | |||||
山麻黄 | 0.0509 | 0.0949 | 0.0057 | 5.05229 | 1 | |||||
卫矛 | 0.0879 | 0.0240 | 0.1429 | 8.4956 | 4.5014 | |||||
绣线菊 | 0.0278 | 0.0160 | 0.0229 | 2.2216 | 1.8250 | |||||
悬钩子 | 0.0787 | 0.0572 | 0.2057 | 11.3858 | 4.6284 | |||||
表3 草本层主要物种的重要值和生态位宽度 | ||||||||||
Table 3 Important values and niche width of main species in herb layer | ||||||||||
物种名 | 相对密度 | 相对盖度 | 相对频度 | 重要值 | 生态位宽度 | |||||
Species name | Relative abundance | Relative coverage | Relative frequency | Important value | Niche breadth | |||||
禾本科 | 0.0281 | 0.0576 | 0.08889 | 5.8205 | 1.9423 | |||||
蕨 | 0.1142 | 0.1371 | 0.08889 | 11.3422 | 3.9766 | |||||
茜草 | 0.0066 | 0.0143 | 0.05 | 2.3655 | 2.8072 | |||||
苔草 | 0.4669 | 0.3646 | 0.3556 | 39.5667 | 3.1576 | |||||
唐松草 | 0.0132 | 0.0306 | 0.05 | 3.1269 | 1.7888 | |||||
崖棕 | 0.1126 | 0.1497 | 0.1389 | 13.372 | 4.3349 | |||||
白茅 | 0.1001 | 0.0616 | 0.0056 | 5.6048 | 1 | |||||
2.2种间联结性
2.2.1物种总体联结性
根据计算得到的重要值,从所有树种中挑选出的9种重要值大于1的乔木物种(见表1)。对这1种主要树种进行整体的联结性计算,得出VR=1.658,VR>1,这些物种之间的整体联结性表现为正,但根据统计量的检验结果W=N×(VR)得出W=14.92,数值落在Χ2(N)0.95<W<Χ2(N)0.05这个区间内,说明该群落中主要树种之间的联结性不够显著,乔木层物种间的关联性并不强烈。
灌木层中的物种选取重要值较大的18种物种(见表二)进行分析,其整体联结性计算结果VR=1.28,VR>1,灌木层总体的联结性是为正的。带入公式W=N×(VR)
表4 乔木层物种间的 Pianka 生态位重叠指数 | |||||||||
Table 4. Peanka niche overlap index among tree species | |||||||||
油松 | 0.58 |
|
|
|
|
|
|
|
|
锐齿栎 | 0.56 | 0.99 |
|
|
|
|
|
|
|
漆树 | 0.77 | 0.76 | 0.69 |
|
|
|
|
|
|
麻栎 | 0 | 0.56 | 0.59 | 0 |
|
|
|
|
|
梾木 | 0 | 0.77 | 0.77 | 0.39 | 0.63 |
|
|
|
|
苦木 | 0.67 | 0.57 | 0.65 | 0.46 | 0.15 | 0.14 |
|
|
|
华山松 | 0.63 | 0.51 | 0.40 | 0.73 | 0 | 0.09 | 0.08 |
|
|
华北落叶松 | 0 | 0.34 | 0.26 | 0.44 | 0 | 0.26 | 0 | 0.37 |
|
鹅耳栎 | 0.34 | 0.61 | 0.57 | 0.50 | 0.15 | 0.30 | 0.60 | 0.31 | 0.60 |
| 灯台树 | 油松 | 锐齿栎 | 漆树 | 麻栎 | 梾木 | 苦木 | 华山松 | 华北落叶松 |
得出W=57.6,数值处于Χ2(N)0.95<W<Χ2(N)0.05的范围内,说明灌木层整体的联结性并不显著。
草本植物由于物种过少不做整体联结性的计算。
2.2.2种对间的联结性
21种木本种物组成的231个种对种中表现出负联结性的一共134对,占总对数的约58%,其中,联结系数大于0.5的(存在显著正联结性)23对,占整体的9.95%,联结系数小于-0.5的(存在显著负联结性)一共25对,约占整体的10.82%,存在显著联结关系的种对占整体种对的20.78%。在25对显著负联结的关系中,有14对都是发生在乔木和灌木中,只有11各发生在同一层次的两个植物身上,而23对显著正联结的种也有14个发生在同一层次的植物中,只有个9发生在不同层次的物种间,草本物种的21个种对中呈正联结性的有9对,其余12对为负联结,正负比值为0.75。其中出现显著联结性的只有1个种对,就是白茅和苔草。
表5灌木层层物种间的 Pianka 生态位重叠指数 | ||||||||||
Table 5. Pianka niche overlap index among shrub species | ||||||||||
绣线菊 | 0.78 |
|
|
|
|
|
|
|
|
|
卫矛 | 0.56 | 0.43 |
|
|
|
|
|
|
|
|
山麻黄 | 0.43 | 0.00 | 0.36 |
|
|
|
|
|
|
|
忍冬 | 0.32 | 0.00 | 0.77 | 0.12 |
|
|
|
|
|
|
桦叶荚蒾 | 0.88 | 0.74 | 0.45 | 0.55 | 0.06 |
|
|
|
|
|
胡枝子 | 0.25 | 0.11 | 0.51 | 0.00 | 0.62 | 0.00 |
|
|
|
|
胡颓子 | 0.26 | 0.00 | 0.00 | 0.00 | 0.25 | 0.00 | 0.76 |
|
|
|
勾儿茶 | 0.22 | 0.47 | 0.36 | 0.00 | 0.00 | 0.00 | 0.25 | 0.00 |
|
|
白檀 | 0.82 | 0.85 | 0.63 | 0.39 | 0.13 | 0.72 | 0.17 | 0.00 | 0.68 |
|
菝葜 | 0.52 | 0.45 | 0.38 | 0.00 | 0.32 | 0.28 | 0.37 | 0.39 | 0.33 | 0.48 |
| 悬钩子 | 绣线菊 | 卫矛 | 山麻黄 | 忍冬 | 桦叶荚蒾 | 胡枝子 | 胡颓子 | 勾儿茶 | 白檀 |
结合生态位重叠指数分析,乔木和灌木一共36对生态位重叠指数较大的种对在种间联结上也存在不同程度的正负联结,但联结性较为显著的只有9对,分别是白檀-绣线菊、白檀-卫矛、白檀-桦叶荚蒾,灯台树-苦木、鹅耳栎-油松、鹅耳栎-锐齿栎、苦木-油松、锐齿栎-漆树、锐齿栎-油松,其中,除漆树-锐齿栎种对间存在负联结性,其余均为正联结关系。而在草本层中有两对,分别是禾本科-唐松草和崖棕-蕨,且都为正联结。
表6 草本层物种间的 Pianka 生态位重叠指数 | |||||||||||||
Table 6. Pianka niche overlap index among shrub species | |||||||||||||
崖棕 | 0.15 |
|
|
|
|
| |||||||
唐松草 | 0.00 | 0.17 |
|
|
|
| |||||||
苔草 | 0.18 | 0.22 | 0.92 |
|
|
| |||||||
茜草 | 0.36 | 0.39 | 0.08 | 0.18 |
|
| |||||||
蕨 | 0.00 | 0.65 | 0.20 | 0.18 | 0.30 |
| |||||||
禾本科 | 0.00 | 0.12 | 0.93 | 0.97 | 0.00 | 0.15 | |||||||
| 白茅 | 崖棕 | 唐松草 | 苔草 | 茜草 | 蕨 | |||||||
表7 草本植物种间联结系数表 | |||||||||||||
Table 7 The coefficient of interspecific association of herbaceous plants | |||||||||||||
禾本科 | -0.23 |
|
|
|
|
| |||||||
蕨 | 0.16 | -0.41 |
|
|
|
| |||||||
茜草 | 0.27 | -0.42 | 0.10 |
|
|
| |||||||
苔草 | 1.00 | -0.22 | 0.12 | 0.30 |
|
| |||||||
唐松草 | -0.24 | 0.76 | 0.04 | -0.19 | -0.27 |
| |||||||
崖棕 | -0.03 | -0.44 | 0.53 | -0.06 | -0.08 | -0.01 | |||||||
| 白茅 | 禾本科 | 蕨 | 茜草 | 苔草 | 唐松草 | |||||||

图1 木本植物种间联结系数图
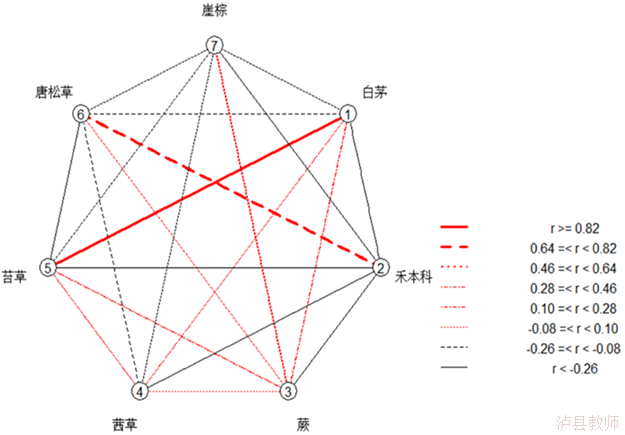
图2 草本植物种间联结系数图
表8 木本植物种间联结系数表 | ||||||||||||||||||||
Table8 The interspecific association coefficient of woody plants | ||||||||||||||||||||
白檀 | 0.26 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
灯台树 | -0.04 | -0.44 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
鹅耳栎 | -0.49 | -0.38 | -0.28 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
勾儿茶 | 0.29 | 0.38 | 0.17 | -0.14 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
胡颓子 | 0.29 | -0.38 | -0.28 | -0.14 | -0.14 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
胡枝子 | 0.07 | -0.26 | 0.34 | -0.29 | 0.49 | 0.49 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
华北落叶松 | 0.29 | 0.38 | -0.28 | -0.14 | -0.14 | -0.14 | -0.29 |
|
|
|
|
|
|
|
|
|
|
|
|
|
华山松 | 0.43 | 0.47 | 0.14 | -0.21 | 0.94 | -0.21 | 0.36 | 0.18 |
|
|
|
|
|
|
|
|
|
|
|
|
桦叶荚蒾 | -0.15 | 0.58 | -0.42 | -0.22 | -0.22 | -0.22 | -0.45 | 0.65 | -0.02 |
|
|
|
|
|
|
|
|
|
|
|
苦木 | -0.58 | -0.75 | 0.62 | 0.47 | -0.28 | -0.28 | 0.06 | -0.28 | -0.35 | -0.43 |
|
|
|
|
|
|
|
|
|
|
梾木 | 0.16 | 0.50 | -0.57 | -0.19 | -0.29 | 0.03 | -0.38 | -0.08 | -0.36 | 0.12 | -0.51 |
|
|
|
|
|
|
|
|
|
麻栎 | 0.03 | -0.38 | -0.43 | 0.09 | -0.22 | 0.94 | 0.33 | -0.22 | -0.33 | -0.10 | -0.24 | 0.04 |
|
|
|
|
|
|
|
|
漆树 | 0.49 | 0.38 | 0.28 | -0.43 | 0.14 | -0.43 | -0.10 | 0.71 | 0.40 | 0.22 | -0.09 | -0.08 | -0.67 |
|
|
|
|
|
|
|
忍冬 | -0.60 | -0.26 | -0.34 | 0.29 | -0.49 | 0.29 | 0.07 | -0.49 | -0.71 | -0.15 | 0.19 | 0.46 | 0.46 | -0.68 |
|
|
|
|
|
|
锐齿栎 | -0.45 | -0.35 | -0.44 | 0.95 | -0.36 | -0.02 | -0.45 | -0.17 | -0.44 | -0.15 | 0.34 | 0.06 | 0.23 | -0.53 | 0.45 |
|
|
|
|
|
山麻黄 | -0.49 | 0.38 | -0.28 | -0.14 | -0.14 | -0.14 | -0.29 | -0.14 | -0.21 | 0.65 | -0.28 | 0.24 | 0.09 | -0.43 | 0.29 | -0.02 |
|
|
|
|
卫矛 | -0.07 | 0.77 | -0.04 | -0.49 | 0.29 | -0.49 | 0.07 | 0.29 | 0.34 | 0.45 | -0.32 | 0.31 | -0.55 | 0.49 | -0.07 | -0.55 | 0.29 |
|
|
|
绣线菊 | 0.45 | 0.58 | -0.08 | -0.22 | 0.65 | -0.22 | 0.15 | 0.65 | 0.85 | 0.33 | -0.43 | -0.29 | -0.34 | 0.65 | -0.75 | -0.41 | -0.22 | 0.45 |
|
|
悬钩子 | -0.28 | 0.42 | -0.65 | 0.14 | 0.14 | 0.14 | -0.13 | -0.36 | -0.03 | 0.28 | -0.51 | 0.46 | 0.38 | -0.68 | 0.46 | 0.26 | 0.73 | 0.13 | -0.17 |
|
油松 | -0.24 | -0.49 | 0.09 | 0.68 | -0.18 | -0.15 | -0.18 | -0.54 | -0.35 | -0.72 | 0.57 | 0.14 | -0.07 | -0.34 | 0.40 | 0.69 | -0.40 | -0.46 | -0.55 | -0.08 |
| 菝葜 | 白檀 | 灯台树 | 鹅耳栎 | 勾儿茶 | 胡颓子 | 胡枝子 | 华北落叶松 | 华山松 | 桦叶荚蒾 | 苦木 | 梾木 | 麻栎 | 漆树 | 忍冬 | 锐齿栎 | 山麻黄 | 卫矛 | 绣线菊 | 悬钩子 |
3 讨论
物种的生态位宽度大小反映着物种在空间资源上利用的综合能力,一般来讲,生态位宽度越大说明该物种利用资源的能力越强,在环境中的适应性也就越强,在群落中的存在更为普遍,也就是我们俗称的优势种[12]。本研究中,乔木层的油松和锐齿栎作为乔木层重要值最大的两个物种也占据着乔木层中最宽广的生态位,是当之无愧的优势种,而灌木层和草本植物层中物种的最大重要值和最高的生态位宽度总数无法很好地匹配,这与杨倩,李宁云等人在大山包湿地的草本植被中调查结果一致[13]。本地区灌丛和草本层发生这种情况的原因可能是林下植物受乔木层盖度的影响,光线成为主要的调节因子,某些植物的适宜生存环境范围狭窄,因此在只在个别样方内形成优势种,而体现出较大的重要值和较小的生态位宽度。
群落中植物能够和平共处其基本条件之一就是要产生生态位的分化[14]。物种间的生态位重叠指数则体现着物种在对同一资源的利用上的一致性,生态位重叠指数越高说明两个物种在对该资源的使用上存在较高的一致性[15],也就是说物种生态位重叠指数越高,它们的生态位分化程度越低,两者应该存在较强的竞争,但分析本文得到的数据来看,在所有生态位重叠指数较高的种对之中,有三分之二都表现为正联结,四分之一存在明显的联结现象的种对几乎全为正联结的关系(只有一对是负联结关系),且这些出现明显正联结关系的物种在各自的群落都有较高的重要值,比如油松和锐齿栎,两者有0.98的生态位重叠指数,却有着显著的正联结性,而两者又都是这个群落的优势种,这可能说明这个群落的优势种已经逐渐固定,对于优势种来说资源已经得到了较合理的分配,在漫长的竞争中已经形成了彼此独特的位置。群落正在朝着顶级群落的方向走去。因此本文更为支持群落演替到越稳定的阶段种间联结性将偏向正联结的观点[16]。同一群落的优势种往往对环境因子的要求较为一致因此有较大的生态位重叠,但它们对环境的适应能力势均力敌,在经过漫长的演化之后彼此资源能合理分割,最终在群落顶级阶段时能够稳定地共存。
总体上来讲,该地区木本植物优势种之间普遍存在的正联结性足以说明该群落具有一定的稳定性[17],但所有种对中种间联结关系明显负值多于正值,且对群落植物整体联结性的计算表明该群落并未表现出显著的联结性,可以证明除了群落中相对优势的物种外,其他物种依旧存在一定的不稳定性,尤其是一些优势度较低的物种就很有可能在竞争的过程中被淘汰。该群落处于群落演替的中后期。
植物种间的相互作用在大自然中是普遍存在的,且其实际作用只有在一定的范围内才会产生,而目前的大部分计算种间联结性的方法只能通过在样方中的对植物的调查和分析,此时样方的大小和数目就成了种间联结性最终分析结果的重要影响因子,而到底多大的样方多少样方数才是最合适的似乎很难确定[18],且运用这样的数学公式模型最终只能得到物种间的共存性的结果,并不能具体反映物种之间到底以何种方式共存或者互斥[19],实验存在误差和偶然,模型更是难以还原群落里复杂的生境,因此想要更加深入的了解群落的种间关系不能只看联结性,还需要与其他研究方式结合。
本文尝试把乔木和灌木两种不同层的植物放在一起分析相关性,发现出现显著相关性的种对十分有意思,正联结性总是存在于同层次的植物种,负联结性总是在不同层次的植物之间不期而遇,到底是什么原因造成这种差别,或者这到底是一个真正存在的差别还是由于低频样方带来的偶然性?想要得到答案还有待更深层次的调查。
结论
本论文研究的区域中油松和和锐齿栎是十分明显的优势种物种,占据群落内较大的重要值和较宽的生态位宽度,且与多种植物形成正联结关系,有十分稳定的地位,对于其余重要值较大的物种来说大部分物种也能在有生态位重叠的同时与其他物种形成较明显的正联结性,群落的共存潜力较大。同时群落中的物种之间也存在一定的负联结作用,可以推测群落中除优势种之外其余物种依旧有较强的竞争,生态位分化不够成熟,群落处于演替的中后期。对于优势种来说它们适应性强,有较宽的生态位宽度,能够在一个群落中稳定地存在,目前群落已逐步稳定,但森林内部还处于不是很稳定的状态。油松是人们为了提高森林郁闭度进行人为撒种形成的种群,却能在当地形成地位稳定的优势种,这说明选择的物种只要适宜当地的生物和自然环境,人类是可以通过一些方法使自然群落向着较好的方向发展的,因此我们在保护或者需要重建某个生态系统时,可以提前调查当地的生境,选择适宜的物种帮助群落发展,同时也要保护当地的优势种的稳定性,当地原有的优势物种已经适应了当地的环境,保护好当地优势种有利于保护一些需要在特定环境下才能生存的物种的繁衍,因此应加强对当地松栎林的保护。
参考文献:
[1]李远杰,张玲.物种间关系网络结构和形成机制的研究进展[J].绿色科技,2018(14):10-13+19.
[2]Ruben Heleno, Cristina Garcia, Pedro Jordano, Anna Traveset, José Maria Gómez, Nico Blüthgen, Jane Memmott, Mari Moora, Jorge Cerdeira, Susana Rodríguez-Echeverría, Helena Freitas, Jens M. Olesen. Ecological networks: delving into the architecture of biodiversity[J]. Biology Letters,2014,10(1).20131000
[3]史浩伯,陈亚宁,李卫红,孙桂丽.塔里木河下游植被种间关系与稳定性分析[J].干旱区研究,2020,37(01):220-226.
[4]徐满厚,刘敏,翟大彤,刘彤.植物种间联结研究内容与方法评述[J].生态学报,2016,36(24):8224-8233.
[5]张先平,王孟本,张伟锋,肖扬.庞泉沟国家自然保护区森林群落木本植物种间关系的分析[J].植物研究,2007(03):350-355+371.
[6]李晶晶. 秦岭中段南坡林区植被优势种能量结构特征研究[D].西北农林科技大学,2013.
[7]孙儒泳,李庆芬,牛翠娟,等.基础生态学[M].北京:高 等教育出版社,2002.
[8] Levins R.Evolution in Changing Environments: Some Theoretical Explorations. Princeton: Princeton University Press,1968.
[9] Pianka E R. The structure of lizard communities. Annual Review of Ecology and Systematics,1973,4: 53-74.
[10]Dolph Schluter. A Variance Test for Detecting Species Associations, with Some Example Applications[J]. John Wiley & Sons, Ltd,1984,65(3) :998-1005
[11]刘润,红,陈乐,涂洪润,梁士楚,姜勇,李月娟,黄冬柳,农娟丽.桂林岩溶石山青冈群落灌木层主要物种生态位与种间联结[J/OL].生态学报,2020(06):1-15[2020-04-20]
[12]刘菊红,王忠武,韩国栋.重度放牧对荒漠草原主要植物种间关系及群落稳定性的影响[J].生态学杂志,2019,38(09):2595-2602.
[13]杨倩,李宁云,陈丽,李杰,闫凯,赵子娇.大山包湿地植被群落数量分类及主要种生态位特征研究[J].西部林业科学,2020,49(02):36-42.
[14]史浩伯,陈亚宁,李卫红,孙桂丽.塔里木河下游植被种间关系与稳定性分析[J].干旱区研究,2020,37(01):220-226.
[15]彭舜磊,陈昌东,李彦娇,刘丹丹,赵干卿.白龟湖国家湿地公园植物群落数量分类及优势植物生态位分析[J].湿地科学,2016,14(05):619-627.
[16]杜道林,刘玉成,苏洁.四川缙云山大头茶与森林优势种群间联结性研究[J].西南师范大学学报(自然科学版),1995(01):66-72.
[17]赵峰.莽山南方铁杉群落种间关系研究[J].中国农学通报,2011,27(31):68-72.
[18]刘萍萍,程积民.植物种间联结关系的研究[J].水土保持研究,2000(02):179-184.
[19]李德志,秦艾丽,杨茂林,刘鹏飞.天然次生林群落中主要树木种群间联结关系的研究[J].植物生态学报,1996(03):263-271.
致 谢
在我毕业论文的调查、研究和撰写期间遇到过很多困难,但是最终都被克服了,这离不开我周围的人对我的帮助,首先要感谢康迪老师给予了我莫大的鼓励和支持,耐心引导我论文设计的走向,数据分析的思路,加深我对研究方法的理解,同时细致地指出我的论文中存在的不足之处,没有康迪老师的帮助我很难顺利完成论文,其次要感谢我的好友叶宇给予我的耐心解答,她是一位很优秀的同学,不仅品学兼优还乐于助人,很感谢在写论文的过程中有她的陪伴,此外我还要感谢我的家人在这段时间对我的关心和照顾,让我能够一心一意投入到论文的写作中,让我感受到家的温暖,最后我要感谢我的学校对本科毕业生的关心。感谢我的学校,我的老师们,朋友们,亲人们,感谢一切对我提供过关心和照顾的人们,在此致敬以上全体!
账号+密码登录
手机+密码登录
微信扫码登录
还没有账号?
立即注册